Si no se regenera NAD⁺ con suficiente rapidez, la glucólisis se ralentiza o incluso se detiene, aunque haya glucosa disponible. La conversión de piruvato en lactato permite reciclar ese NADH de nuevo a NAD⁺, lo que mantiene activa la producción rápida de energía cuando la demanda es alta.
Este sistema:
- Produce ATP más rápido que el metabolismo oxidativo.
- No depende directamente del aporte inmediato de oxígeno.
- Genera lactato como intermediario metabólico.
- Es dominante en esfuerzos de intensidad media-alta y en cambios de ritmo.
Aquí aparece un concepto clave para entender el rendimiento: el lactato no es un “residuo” del metabolismo ni el responsable directo de la fatiga, como durante años se pensó. En realidad, es un metabolito útil que se produce cuando el músculo genera energía rápidamente a partir de la glucosa.
Lejos de ser un problema, el lactato puede reutilizarse como combustible por otras fibras musculares, por el corazón o incluso por el propio músculo que lo produjo. Cuando su concentración aumenta, lo que realmente está indicando es que la producción de energía es muy alta y que, de forma temporal, la velocidad a la que se produce supera la capacidad de las mitocondrias para oxidarlo. No es un fallo del sistema, sino una señal de que el metabolismo está trabajando a gran intensidad. En modelos actuales, como la teoría de la lactate shuttle descrita por George A. Brooks, el lactato se entiende como un intermediario dinámico dentro de una red energética integrada, cuya función se desarrolla con mayor profundidad en artículos posteriores.
 Metabolismo oxidativo (mitocondrial)
Metabolismo oxidativo (mitocondrial)
Es el sistema más eficiente y el más determinante en deportes de resistencia, ya que permite producir grandes cantidades de energía de forma sostenida durante horas, siempre que la intensidad sea compatible con la capacidad oxidativa del músculo.
Cuando el piruvato entra en la mitocondria, se transforma en acetil-CoA y accede al ciclo de Krebs. En términos sencillos, la mitocondria actúa como una “central energética” donde los nutrientes se terminan de procesar para extraer la máxima cantidad de energía posible. Los electrones liberados durante estas reacciones pasan a la cadena respiratoria, que funciona como una especie de “turbina” bioquímica donde, utilizando oxígeno, se fabrica ATP de forma muy eficiente.
Además de la glucosa, los ácidos grasos también pueden convertirse en acetil-CoA mediante β-oxidación y alimentar este mismo sistema. Esto explica por qué, en intensidades bajas y medias, el organismo puede sostener el esfuerzo durante mucho tiempo utilizando principalmente grasa como combustible, ahorrando glucógeno y retrasando la aparición de la fatiga.
Este sistema:
- Necesita oxígeno.
- Produce grandes cantidades de ATP.
- Es más lento en activarse que los sistemas anaeróbicos.
- Es altamente eficiente a largo plazo.
- Genera menos lactato cuando está bien desarrollado.
Estas adaptaciones no ocurren de forma espontánea. El ejercicio activa sensores energéticos intracelulares que regulan la expresión de genes relacionados con la biogénesis mitocondrial. Entre ellos destaca PGC-1α, considerado el principal regulador de la adaptación oxidativa inducida por el entrenamiento. Cada sesión no sólo genera fatiga transitoria, sino una señal molecular que remodela progresivamente la maquinaria energética del músculo.
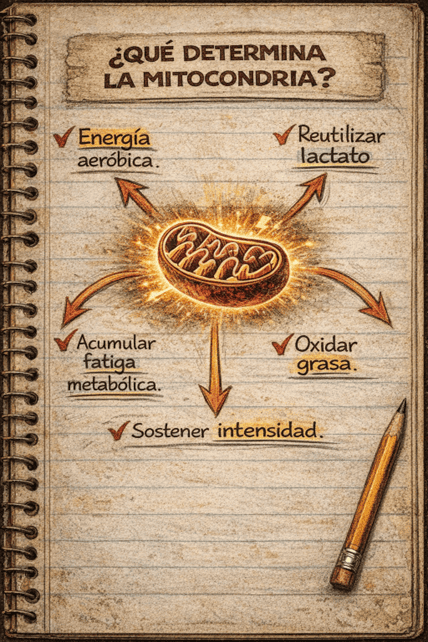
Desde un punto de vista aplicado al entrenamiento, cuanto mayor sea tu capacidad mitocondrial (más mitocondrias y más eficientes), mayor será tu capacidad para producir energía de forma estable, utilizar mejor las grasas, reciclar el lactato producido en otras fibras o tejidos y sostener intensidades relativamente altas con menor acumulación de fatiga metabólica.
En la práctica, “entrenar el sistema aeróbico” es entrenar la maquinaria que te permite ir más rápido durante más tiempo, con menor coste energético por unidad de velocidad o potencia.