Entrenamiento en zona 3 , 4 y 5:
del puente metabólico a la potencia mitocondrial
Entrenamiento en Zona 3: adaptación inmediata
La intensidad intermedia aumenta la dependencia glucolítica y la producción de lactato, pero sin alcanzar el estrés máximo de la zona 5 ni la señal oxidativa pura de la zona 2.
Produce adaptación, pero desde un punto de vista mitocondrial es menos específica.
Cuando esta zona domina el entrenamiento, puede aumentar la fatiga sin maximizar ni la eficiencia oxidativa ni la capacidad máxima del sistema energético.
Esto ayuda a entender por qué muchos modelos contemporáneos de entrenamiento de resistencia evitan acumular demasiado volumen en esta franja de intensidad.
Comprender esto permite contextualizar el papel del lactato: no representa un límite metabólico rígido, sino una variable dinámica dentro de un sistema energético integrado.
Lactato: el puente metabólico (George Brooks)
Para entender el papel del lactato en el rendimiento primero hay que comprender qué sucede en cada contracción muscular.
Cada vez que una fibra muscular se contrae necesita energía en forma de ATP. Una de las vías más rápidas para obtenerlo es la glucólisis, proceso mediante el cual la glucosa se transforma en piruvato generando ATP en el citoplasma.
El piruvato es uno de los intermediarios más importantes del metabolismo energético.
♦ Qué ocurre con el piruvato dentro de la fibra muscular
La mayor parte del piruvato producido en una fibra muscular se utiliza dentro de esa misma fibra, especialmente si la capacidad oxidativa de la célula es suficiente.
Es decir, cuando la intensidad del ejercicio permite que el metabolismo aeróbico funcione con normalidad, el piruvato entra directamente en la mitocondria de esa misma célula para ser oxidado.
Cuando la glucosa se metaboliza mediante glucólisis en el citosol se generan dos moléculas de piruvato. A partir de ahí pueden ocurrir dos destinos metabólicos principales.
- Entrada en la mitocondria (vía predominante en entorno aeróbico)
- El piruvato es transportado al interior de la mitocondria mediante el transportador MPC (mitochondrial pyruvate carrier).
- Dentro de la mitocondria es convertido en acetil-CoA por el complejo piruvato deshidrogenasa (PDH).
- A partir de ahí entra en el ciclo de Krebs, generando NADH y FADH₂ que posteriormente alimentan la cadena respiratoria, permitiendo producir ATP mediante fosforilación oxidativa.
- Este es el destino predominante del piruvato cuando la intensidad del ejercicio es compatible con la capacidad oxidativa del músculo.
- Conversión a lactato (cuando la capacidad oxidativa se satura)
- Cuando la intensidad aumenta, la velocidad de producción de piruvato puede superar la capacidad de la mitocondria para oxidarlo.
- En ese momento el piruvato se convierte en lactato mediante la enzima lactato deshidrogenasa (LDH).
- Esta reacción permite regenerar NAD⁺, molécula imprescindible para que la glucólisis pueda continuar produciendo ATP a alta velocidad.
- Por tanto, la formación de lactato no es un error metabólico.
Es un mecanismo que permite mantener activa la producción rápida de energía.
♦ ¿Puede el piruvato viajar a otra fibra muscular?
En condiciones fisiológicas ésto es muy poco probable.
El piruvato rara vez se transporta entre células musculares de forma significativa por varias razones:
- Es químicamente menos estable que el lactato.
- Su concentración intracelular se mantiene muy baja.
- Las membranas celulares no están diseñadas para exportarlo de forma eficiente.
Por este motivo el organismo utiliza un mecanismo más eficaz: convertir el piruvato en lactato para poder transportarlo.
♦ Por qué el lactato sí puede viajar
El lactato se transporta fácilmente entre células gracias a los transportadores MCT (monocarboxylate transporters).
Los dos más importantes en músculo esquelético son:
⇒ MCT4
- Predomina en fibras tipo II (rápidas y glucolíticas).
- Su función principal es exportar lactato desde las fibras que lo producen.
⇒ MCT1
- Predomina en fibras tipo I (lentas y oxidativas).
- Su función es introducir lactato dentro de la célula para oxidarlo.
Una vez dentro de una fibra oxidativa, el lactato puede:
- Convertirse nuevamente en piruvato
- Entrar en la mitocondria
- Oxidarse para producir ATP
Este sistema de redistribución energética es lo que George Brooks denominó lactate shuttle.
La lanzadera del lactato o lactate shuttle
Durante décadas se pensó que el lactato era un residuo responsable de la fatiga.
Sin embargo, la teoría de la lactate shuttle cambió completamente esa visión.
El lactato es un intermediario metabólico que conecta el metabolismo glucolítico con el metabolismo oxidativo.
Su dinámica básica es la siguiente:
- Las fibras tipo II producen lactato cuando la glucólisis es elevada.
- Estas fibras lo exportan mediante MCT4.
- Las fibras tipo I lo captan mediante MCT1.
- El lactato se reconvierte en piruvato.
- El piruvato entra en la mitocondria y se oxida.
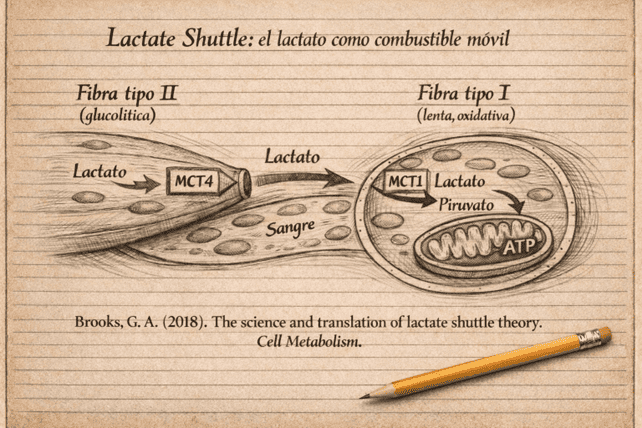
♦ Destino sistémico del lactato
El lactato no sólo circula entre fibras musculares. También puede ser utilizado por otros tejidos del organismo.
Corazón: El miocardio utiliza lactato como uno de sus combustibles preferentes.
Hígado: Puede convertir lactato en glucosa mediante el ciclo de Cori.
Cerebro: En determinadas condiciones también puede utilizar lactato como fuente energética.
Esto demuestra que el lactato actúa como una molécula de redistribución energética sistémica.
¿El lactato causa la acidez muscular?
Durante años se afirmó que el lactato era responsable de la acidosis muscular.
Sin embargo, investigaciones posteriores —como el trabajo de Robergs et al. (2004)— demostraron que la acidez muscular no es causada por el lactato, sino por la acumulación de protones (H⁺) derivados del metabolismo energético.
De hecho, la conversión de piruvato en lactato consume protones, contribuyendo parcialmente a amortiguar la acidez.
El lactato no es el problema.
La acumulación de protones es la verdadera responsable de la caída del pH muscular.
♦ El bicarbonato: principal sistema tampón extracelular
Cuando durante el ejercicio intenso aumenta la concentración de protones (H⁺), el organismo activa varios sistemas tampón para limitar la caída del pH. El principal en el espacio extracelular es el bicarbonato, que reacciona con los protones y contribuye a su transformación en CO₂ y agua, permitiendo que parte de esa carga ácida se elimine a través de la respiración:
H⁺ + HCO₃⁻ ⇄ H₂CO₃ ⇄ CO₂ + H₂O
Pero no actúa solo: también contribuyen las proteínas, la hemoglobina y los fosfatos, especialmente en la sangre y en el medio intracelular. En conjunto, estos sistemas no eliminan por sí mismos la carga ácida, pero sí amortiguan su impacto y ayudan a mantener un entorno químico compatible con la contracción muscular y el rendimiento.
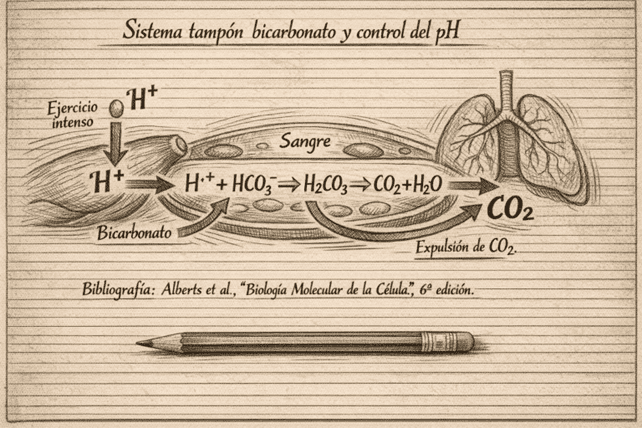
Por eso, cuando entrenamos cerca o por encima del umbral, aumenta la ventilación pulmonar, se incrementa la expulsión de CO₂ y el organismo consigue estabilizar parcialmente el pH. En términos prácticos, cuanto mejor entrenado está un deportista, mayor suele ser su capacidad de tolerar y amortiguar cargas elevadas de protones, retrasando la fatiga asociada al esfuerzo intenso.
Este mecanismo ayuda a explicar por qué la suplementación con bicarbonato sódico puede mejorar el rendimiento en esfuerzos de alta intensidad de aproximadamente 1 a 10 minutos: aumenta la capacidad tampón extracelular y favorece la salida de H⁺ desde el músculo hacia el espacio extracelular.
Este equilibrio entre producción de protones, reutilización del lactato y capacidad tampón es precisamente lo que determina en gran medida la tolerancia al esfuerzo intenso. Y es en las zonas 4–5 donde este sistema alcanza una de sus máximas exigencias fisiológicas.
Zona 4–5: potencia mitocondrial y mitohormesis
En intensidades cercanas o superiores al umbral:
- Aumenta notablemente la glucólisis
- Aumenta la producción de lactato
- Se incrementa la generación de ROS mitocondriales
Este aumento controlado de especies reactivas de oxígeno actúa como señal adaptativa, fenómeno conocido como mitohormesis.
Este estrés metabólico estimula:
- Mayor expresión de PGC-1α
- Aumento de la capacidad oxidativa máxima
- Mejora del VO₂max
- Optimización de la lanzadera del lactato
En estas intensidades no se mejora tanto la eficiencia energética como la capacidad máxima de producir energía.
Producción vs aclaramiento de lactato: el verdadero equilibrio metabólico
La concentración de lactato en sangre depende del equilibrio entre dos procesos simultáneos:
♦ Producción de lactato
Principalmente en fibras tipo II cuando la glucólisis genera piruvato rápidamente.
♦ Aclaramiento de lactato
Principalmente en fibras tipo I, corazón, hígado y otros tejidos capaces de oxidarlo.
Mientras el organismo puede reutilizar el lactato al mismo ritmo al que se produce, su concentración permanece relativamente estable.
Este equilibrio depende en gran medida de la capacidad mitocondrial del sistema.
Cuando la red oxidativa es potente:
- Más piruvato entra en la mitocondria
- Más lactato se reutiliza
- La concentración sanguínea se mantiene baja
- El esfuerzo es metabólicamente estable
Cuando la producción supera la capacidad de aclaramiento:
- El lactato comienza a acumularse
- Aumenta la participación glucolítica
- Aparece mayor estrés metabólico
Este desequilibrio progresivo es lo que define fisiológicamente los umbrales metabólicos del ejercicio.
Implicación para el entrenamiento
Desde esta perspectiva, el objetivo del entrenamiento no es evitar el lactato, sino aumentar la capacidad del organismo para gestionarlo.
Esto se consigue mediante estímulos complementarios:
Zona 2
- Aumenta la capacidad de oxidar lactato
- Mejora la eficiencia mitocondrial
- Aumenta la expresión de MCT1
Zonas 4–5
- Aumentan el flujo glucolítico
- Incrementan la producción de lactato
- Aumentan la expresión de MCT4
Cuando ambos sistemas se desarrollan de forma equilibrada, el organismo puede producir grandes cantidades de energía sin que el lactato se convierta en un factor limitante inmediato.
Por eso el lactato no es simplemente un marcador de fatiga.
Es una señal del equilibrio dinámico entre producción y reutilización de energía dentro del sistema metabólico

J.R.Celada (Col 56.782).
Referencias científicas
- López Chicharro, J., & Legido Arce, J. C. (2008). Umbral anaeróbico y entrenamiento. McGraw-Hill Interamericana.
- López Chicharro, J., & Fernández Vaquero, A. (2006). Fisiología del ejercicio. Editorial Médica Panamericana.
- Chicharro, J. L., & Vaquero, A. F. (2006). Bases fisiológicas del metabolismo energético durante el ejercicio. Archivos de Medicina del Deporte.
- Calleja-González, J., Mielgo-Ayuso, J., & León-Guereño, P. (2017). Estrategias nutricionales y tamponamiento en ejercicios de alta intensidad. Revista Andaluza de Medicina del Deporte, 10(4), 185-192.
- Brooks, G. A. (2018). The science and translation of lactate shuttle theory. Cell Metabolism, 27(4), 757-785.
- Granata, C., Jamnick, N. A., & Bishop, D. J. (2018). Training-induced changes in mitochondrial content and respiratory function in human skeletal muscle. Sports Medicine.
- Bishop, D. J., Botella, J., & Genders, A. J. (2019). High-intensity exercise and mitochondrial biogenesis: current controversies and future research directions. Physiology,